Pharmacokinetics and Mechanisms of Action Against Gastrointestinal Inflammation, Anxiety, Nausea, (CNS) Cell Damage and Others
This is a comprehensive review of existing scientific work on a non-psychoactive cannabinoid “Cannabigerol”, its pharmacokinetics and pharmacodynamics as well as its mechanisms of action inside the human Endocannabinoid cell signaling system. As it is shown, CBG can inhibit pain signal propagation in the body, inhibit inflammatory response in the immune system, act as a neuroprotector of the central nervous system cells and have a powerful anti-anxiety effect(1), as well as other effects such as anti-bacterial against (MRSA)(2), Streptococcus Mutans, and other anti-biotic resistant bacterial strains by reducing membrane fluidity and causing a consequent increase in membrane permeability(3). Other experiments have demonstrated that CBG is a potential agent against skin conditions like acne, skin inflammation and psoriasis(1), bacterial biofilm (a key factor for contamination of medical devices) favoring the raising of human chronic infections(4).
Cannabigerol (CBG) and its Interaction with Endocannabinoid Cell Signaling System
Cannabigerol (CBG) in it’s acid form (CBGA) is called the “Mother Cannabinoid” because it’s the first cannabinoid that occurs in the cannabis plant, along with various enzymes and other compounds. Synthase reactions between CBGA and enzymes give birth to CBDA, THCA and CBCA. After the cannabis plants are harvested and decarboxylated, the acid forms of these three major phyto-cannabinoids convert into their base forms (CBC, CBD, and Δ9-THC), which are considered more active than their acid forms (Figure 1). During this process, CBG is depleted and found in smaller amounts in the plant post-extraction compared to its original levels. Therefore, to extract a significant amount of CBGA, a specific hemp strain with a high CBGA content is required (See Figure 1 for CBG and CBGA molecules).
Figure 1. Chemical structure of CBG (left) and CBGA (right)
In the presence of inflammatory stimuli in the body, cells of the central nervous system biosynthesize endocannabinoid anandamide (AEA), which is a lipid mediator synthesized ‘on demand’ from cell membrane phospholipids. Once synthesized, AEA activates CB1 and CB2 cannabinoid receptors to elicit a biological response of Dopamine release (see Figure2), after which AEA endocannabinoids are inactivated through re-uptake facilitated by enzymatic degradation by N-acyl ethanolamine-hydrolysing acid amidase (NAAA) and fatty acid amide hydrolase (FAAH) enzymes(5). The CB1 and CB2 cannabinoid receptors are part of the endocannabinoid signaling system which also includes the enzymes that synthesize and degrade endocannabinoids, as well as possible transporters. The molecular mechanisms for regulating lipid-based signaling events such as cannabinoid receptor signaling are not yet completely understood, although significant progress has been made(6,7). Activation of CB1 and CB2 receptors results in some natural inflammatory stimulus suppression(8), but the amplitude and duration of that suppression is regulated by how fast they are broken down, so this anti-nociceptive effect is weak and short-lived (only about 4-5 minutes in vivo)(9). It has been demonstrated that Cannabigerol (CBG) causes inhibition of the reuptake/downregulation/metabolism/degradation of AEA as a result of activation and desensitization of TRPA1, TRPV1, and TRPV2 cellular channels, while also deactivating TRPM8 cellular channel(1). This leads to CB1 and CB2 receptors remaining active for a much longer time, and a longer pain relief(11).
CBG also blocks the voltage-gated sodium channels (Nav 1.1, 1.2, and 1.5) and can reduce the psychotropic effect of THC(12). Both CBG and AEA possess anti-inflammatory activity through a not sufficiently clarified mechanism of action(1).

Figure 2. The endocannabinoid system – consisting of the endocannabinoids and the cannabinoid receptors – regulates nerve cell communication at the synapse, thereby playing a role in a variety of bodily functions. Activation of Dopamine receptors “D2” triggers release if Anandamide (AEA) endocannabinoid, leading to release of Dopamine(10).
COX-mediated anti-inflammatory activity of CBG and CBGA have been studied and both showed inhibition of the enzyme COX-1 and COX-2 activity by > 30% (13).
Analysis of Some of the Existing Evidence of GI Anti-Inflammatory and Antioxidant Effects of Cannabigerol(CBG)
Inflammatory bowel disease (IBD) is an incurable disease, which affects 200 per 100,000 adults in the United States and 400 per 100,000 in the United Kingdom(15). Major subtypes consist of Crohn’s disease and ulcerative colitis(15). A definitive clinical treatment for these chronic relapsing diseases remains elusive, as currently no therapy exists to reverse the clinical pathology without a risk of significant side effects(15). Corticosteroids, 5-ASA agents, anti-TNFα antibodies, and other immunomodulatory drugs have all been shown to induce significant remission in IBD, but are associated with bone marrow suppression, opportunistic infection, infusion reactions, and malignancy secondary to immunosuppression(15). Hence, it is required to develop new approaches with fewer side effects for the treatment of IBD(16).
Anti-inflammatory and anti-oxidant activities were investigated mainly through pre-clinical research. In laboratory investigations, CBG has been shown to have anti-inflammatory effects, and a subgroup analysis suggested that in experimental colitis CBG caused the largest reduction in inflammation among other well known cannabinoids tested(15). Presented below is a review of the investigation of the effect of CBG on Ulcerative Colitis (UC), which is characterized by abdominal pain, diarrhea, bleeding and malabsorption(16). Inflammation was assessed by evaluating inflammatory markers/parameters (colon weight/colon length ratio and myeloperoxidase activity), by histological analysis and immunohistochemistry; interleukin-1b, interleukin-10 and interferon-g levels by ELISA, inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) by western blot and RT-PCR; CuZn-superoxide dismutase (SOD) activity by a colorimetric assay(16).
IBD causes a significant increase in colon weight/colon length ratio, a simple and reliable marker of intestinal inflammation/damage and a significant decrease in colon weight/length ratio was achieved after administration of CBG in the concentration range of (1mg/kg – 30mg/kg) in a dose dependent manner (See Figure 3)(16). To confirm the anti-inflammatory activity of CBG we performed histological analysis of Control, IBS and 30mg/kg CBG treated colons (Figure 4)(16). The curative action of CBG was further confirmed by Intestinal Permeability study. FITC-conjugated dextran presence was not detected in the serum of healthy control animals, which is suggestive of intestinal membrane integrity (Fig. 5A). Serum of IBS subjects contained significant FITC-conjugated dextran, indicating disruption of intestinal membrane integrity (Fig. 5A). CBG treatment (30 mg/kg) completely abolished increased intestinal permeability (Fig. 5A)(16).

Figure 3. Colon weight/length ratio (mg/cm) of colons from normal (Control) and IBS in the presence or absence of (1-30mg/kg) of CBG (16).

Figure 4. Histological evaluations of inflamed and non-inflamed colons: effect of CBG). No histological modification was observed in the mucosa and submucosa of Control colon (A); IBS colon (B); treatment with 30mg/ml CBG reduced colon injury by stimulating regeneration of the glands (C (16).

Figure 5. Effect of CBG on intestinal permeability (evaluated as FITC-dextran permeability), myeloperoxidase (MPO, a marker of intestinal inflammation) activity (B) and superoxide dismutase (SOD) activity (C) (16).
MPO activity is considered to be an index of neutrophil infiltration (because MPO is predominantly found in these cells) and it is largely used to quantify intestinal inflammation(16). Colitis was associated with significantly increased neutrophil infiltration, as evaluated by MPO (Fig. 5B)(16). CBG, given after the inflammatory insult at the dose of 30 mg/kg, counteracted increase in MPO activity (Fig. 5B), hence decreased inflammation(16). Colitis also produced a significant decrease in Superoxide Dismutase (SOD) activity (Fig. 5C), while CBG, at the dose of 30 mg/kg counteracted the reduction (Fig. 5C)(16).
IBS also causes significant production of Reactive Oxygen Species (ROS) in the intestinal epithelial cells, but a pre-treatment for 24 h with CBG (0.1–10 mM) reduced ROS formation as measured by the inhibition of DCF fluorescence intensity(16). The effect was significant starting at the concentration of 1 mM (Fig.6), and CBG (up to 10 mM) had no significant cytotoxic effect on colonic epithelial cells after a 24-h exposure.
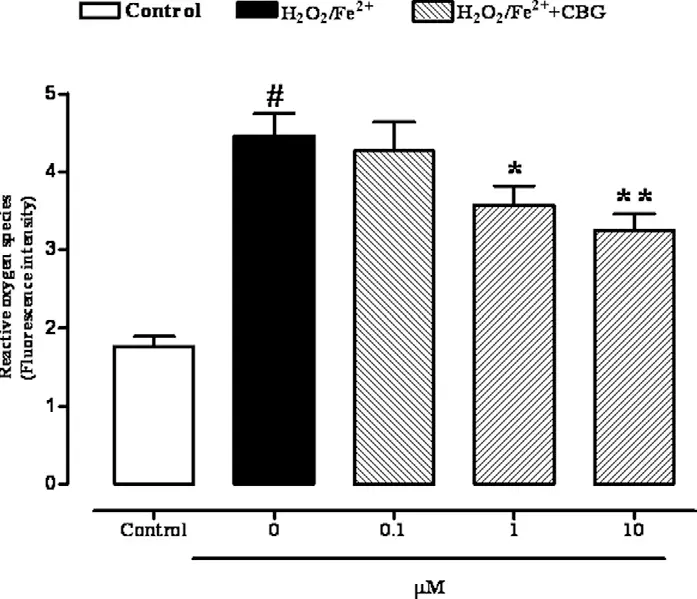
Figure 6. Reactive oxygen species (ROS) production produced by Fenton’s reagent (2 mM H2O2/Fe2+) in Ptk6 null colonic epithelial cells after 24-h exposure to cannabigerol (0.1–10 mM). Results are mean ± SEM of five experiments. #p < 0.001 vs control, *p < 0.05 and **p < 0.01 vs H2O2/Fe2+alone(16).
Discussion
The studies have found that CBG reduced colon weight/colon length ratio of the inflamed colonic tissue, which is considered a reliable and sensitive indicator of the severity and extent of the inflammatory response. CBG was effective when given both before and after the inflammatory insult, suggesting a preventive and a curative (therapeutic) beneficial effect. Significant protective effects were achieved starting from the 1 mg/kg dose (preventive dose) and 5 mg/kg (therapeutic dose). Maximal efficacy was achieved with the 1 mg/kg dose and the30mg/kg dose in the preventative and therapeutic cases respectively. Histological examination showed that CBG 30 mg/kg reduced the signs of colon injury; specifically, in the colon of CBG-treated animals, the glands were regenerating, the oedema in submucosa was reduced and the infiltration of granulocytes into the mucosa and submucosa was decreased. The therapeutic effect of CBG was further demonstrated by its capacity to reverse the increase in intestinal permeability and restored the integrity of intestinal epithelium.
Finally, the evidence shows a possibility that CBG could protect the intestinal mucosa by reducing oxidative stress. SOD activity measurement showed an important antioxidant defense in the gut by eliminating ROS production, a major tissue-destructive force which contributes significantly to the pathogenesis of IBD. These results suggest that the therapeutic effect of CBG could be due, at least in part, to its antioxidant action.
It has been found that by interacting with a CB2 receptor CBG increases cellular antioxidant defense by modulating Superoxide Dismutase (SOD-1) expression, thus inhibiting cell death(17). This indicates that CBG could be useful as a new approach in the care of oxidative stress-related disorders. CBG may effectively work as a free radical scavenger to enhance cellular antioxidant activity through the modulation of pathways such as MAPK kinase and NF-κB translocation and to counteract cell death.
Neuroprotective Effects of CBG
Neuroprotective effects of CBG and CBD have been compared in experiments simulating oxidative stress and neurotoxicity by exposing the assaulted hypothalamic cells to either 1000nM CBD or 1nM CBG, and their influence on the synthesis and release of dopamine (DA), norepinephrine (NE), and serotonin (5-HT) was evaluated, as well as the 3-hydroxykinurenine/kinurenic acid (3-HK/KA) ratio was also determined(18).
3-hydroxykynurenine (3HK) is a metabolite of tryptophan that exhibits cytotoxicity through mechanisms that culminate in apoptosis, or cell death(19). Conversely Kinurenic Acid (KA) is an important bio-active product of tryptophan metabolism, which exhibits well-known neuroprotective effects on mental health disorders(20). So the larger the (3-HK/KA) ratio the larger is neurotoxicity, and vice versa.
To better evaluate the effect of CBD and CBG on the Hypo-E22 hypothalamic cells in their basal state, the detection of the extracellular release of 3-HK and KA was performed by (di Giacomo, Viviana et al.)(18). The ratio 3-HK/KA, a well-known index of neurotoxicity, was considerably reduced following CBD and CBG treatment (Figure 7)(18).

Figure 7. Inhibitory effects induced by 1000nM CBD and 1nM CBG on extracellular 3-hydroxykinurenine/kynurenic acid (3-HK/KA) ratio in hypothalamic Hypo-E22 cells. ANOVA, p<0.001; ** p<0.01, * p<0.05 vs. control group(18).
In parallel, a significant inhibition (p < 0.0001) of Norepinephrine (NE) steady state level was observed when the hypothalami were exposed both to CBD and CBG(18) (Figure 8).

Figure 8. Effects of 1000nM CBD and 1nM CBG on norepinephrine (NE) levels (ng/mg wet tissue) in isolated rat hypothalamus. CBD and CBG inhibited NE levels.
Norepinephrine, or Noradrenalin is a naturally occurring chemical in the body that acts as both a hormone and neurotransmitter (a substance that sends signals between nerve cells), according to the Endocrine Society(21). Stress triggers the release of norepinephrine and adrenaline, which is known as the fight-or-flight response, the body’s emergency response to danger or perceived danger, which causes several changes in the body, including: pupil dilation, pale skin, high blood pressure, rapid irregular heartbeat, excessive sweating, severe headache, nervous feeling and jitters, deeper and faster breathing, burst of energy, burst of blood and oxygen in the muscles according to Cleveland Clinic. However too little norepinephrine can result in: anxiety, depression, attention deficit hyperactivity disorder (ADHD), headaches, memory problems, sleeping problems, low blood pressure (hypotension), low blood sugar (hypoglycemia), changes in blood pressure and heart rate, dopamine beta-hydroxylase deficiency, which is why the noradrenaline level must be carefully regulated and stabilized according to Cleveland Clinic.
Discussion
The protective effects exerted by CBD and CBG on hypothalamic cells challenged with Hydrogen Peroxide (H2O2), as demonstrated by the assay results presented above, are confirmed by the modulation of the release of kynurenine metabolites by the same cells. The 3-HK and KA are key products of the kynurenine pathway, which represents the two main tryptophan degradative systems(22). Additionally, tissue and plasma levels of these two molecules are well known to be related to inflammatory and oxidative stress conditions in both peripheral and central tissues (23,24,25). Specifically, the 3-HK/KA is a reliable marker of neurotoxicity(26), and the findings of reduction of this ratio from Hypo-E22 cells after pharmacological treatment further support the neuroprotective role exerted by both CBD and CBG(18). The CBD modulates hypothalamic neuromodulators, whereas the CBG effect on the same mediators suggests alternative mechanisms, possibly involving peripheral pathways, which is why a combination of CBG and CBD may dose dependently amplify the neuroprotective effects achieved by CBD, or CBG individually.
Anti-Anxiety Effects of CBG
Endocannabinoids have been shown to modulate emotional behaviors(27,28). CBG acts as a serotonin antagonist as demonstrated in in-vitro(29) and in-vivo experiments(30). It is also clearly established (via double-blind placebo-controlled studies)(32) that serotonin receptor agonists have anxiolytic and ant depressive properties, which is why it has been proposed that it can be responsible for the anti-anxiety effects of hemp and cannabis(31).
In other experiments, it has been observed that i.p. CBG administration (intraperitoneal injection) at a dose of 10 mg/kg in mice enhanced the period of time consumed in the central quadrant of the open field test, thus suggesting potential anxiolytic effects. In the same group of experiments, i.p. CBG administration at a dose of 3 mg/kg also produced a light anti-nociceptive effect(33).
Anti Acne, Skin Inflammation and Psoriasis Effects of CBG
The effects of CBG on the expression of keratins, and DNA methylation of keratin 10 gene, have been studied in the human keratinocytes (HaCaT) cellular line, together with DNA methylation and expression of four DNA methyltransferases (DNMT1, 3a, 3b, and 3L). These experiments showed that CBG caused a significant reduction in the expression of the genes investigated by increasing the DNA methylation of the gene for keratin 10. The data obtained from these experiments led the authors to believe that CBG behaves as a transcriptional suppressor, able to control cell proliferation and cell differentiation, being a substance potentially useful for new therapeutic approaches for skin disorders(34).
The effects of CBD, CBG and CBDV were tested in proliferating and differentiated HaCaT cells, and were compared to those of the Endocannabinoid (AEA) as a control(37,38). In a preliminary set of dose-response experiments on K10 gene expression levels (Figure 9), the lowest effective dose of CBD (p < 0.001) and CBG (p < 0.05) was found to be 0.5 µM, whereas CBDV was ineffective up to 1.0 µM, previously found to be the lowest effective dose of AEA(38).

Figure 9. Expression of K10 gene in HaCaT cells. Keratinocytes were induced to differentiate by treatment with TPA plus calcium for 5 days. Differentiated HaCaT cells were treated with 1 μM AEA and different amounts (0.1 – 0.5 – 1.0 μM) of CBD, CBG and CBDV. K10 was detected by quantitative RT-PCR. For the quantitation of gene expression, β-actin was used as housekeeping gene. The results are shown as fold induction over proliferating cells of three independent experiments. Prol, proliferating cells; Ctrl, differentiated cells. ***, p<0.001 vs Prol; ###, p<0.001 vs Ctrl; ##, p<0.01 vs Ctrl; #, p<0.05 vs Ctrl.
By using qRT-PCR analysis, significant reduction of the expression of K10 and TGase5 genes was achieved upon treatment of differentiated HaCaT cells with 0.5 µM CBD (p < 0.001) or CBG (p < 0.05 for K10; p <1 0.001 for TGase5)(34).
On this basis, CBG has been studied as a possible anti-acne drug. Acne is a common skin pathology; however, its more serious expression can deeply weaken the quality of life and, because of social censure, can produce consequent psychological disturbance(35). CBG (10–20 mM) treatment for 24 h significantly reduced lipogenesis stimulated by AEA. These results suggest the possibility that CBG may act as a partial agonist through the same pro-lipogenic signaling pathway on which AEA is active. CBG was also found to suppress inflammation caused by LPS on sebocytes. The results of the above-exposed experiments according to the antiproliferative action of CBG, increase the hypothesis that these compounds may be useful in acne and in other skin pathologies associated with inflammatory characteristics, such as psoriasis(36).
Conclusion
The study of CBG pharmacology shows that this compound shares some characteristics with other phytocannabinoids, but it displays its own characteristic profile, as shown by emerging research(1). Pharmacodynamic research suggests a mechanism of action involving only partial activity of classical cannabinoid receptors, indicating that CBG, as well as CBD, has a multitarget pharmacological action and interacts with a number of endocannabinoid and non-endocannabinoid targets(1). Research, both in vivo and in vitro, shows several pharmacological effects of CBG, such as a dermatological, anti-inflammatory, antioxidant, and anti-anxiety activities. CBG synthetic analogs have been tested with positive effects, particularly in the field of neuroprotection(1). (Figure 10) shows interaction of a synthetic CBG analog with the Endocannabinoid System(1). Promising results obtained with CBG, together with the apparent lack of psychotomimetic THC-like effects, suggest that research on CBG deserves to be deepened as it could be used, alone or in association, for novel therapeutic approaches for several disorders, however, these effects have been proven only through preclinical experiments, and clinical research is needed to confirm these potential activities in humans(1).
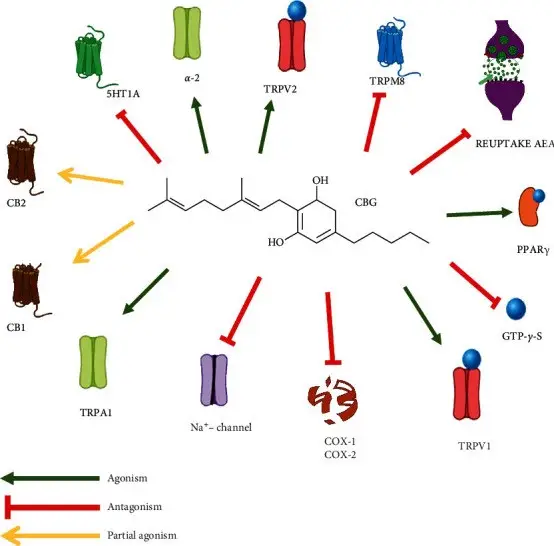
Figure 10. Interaction of a synthetic CBG analog with the Endocannabinoid System.
References
- Fabrizio Calapai, et al. “Pharmacological Aspects and Biological Effects of Cannabigerol and Its Synthetic Derivatives” Evidence Based Complementary Alternative Medicine 2022; 2022: 3336516.
- Appendino G., Gibbons S., Giana A., et al. “Antibacterial cannabinoids from Cannabis sativa: a structure-activity study.” Journal of Natural Products . 2008;71(8):1427–1430.
- Aqawi M., Sionov R. V., Gallily R., Friedman M., Steinberg D. “Anti-bacterial properties of cannabigerol toward Streptococcus mutans.” Frontiers in Microbiology . 2021;12 doi: 10.3389/fmicb.2021.656471.656471.
- Aqawi M., Gallily R., Sionov R. V., Zaks B., Friedman M. Steinberg D. “Cannabigerol prevents quorum sensing and biofilm formation of Vibrio harveyi.”Frontiers in Microbiology . 2020;11:p. 858. doi: 10.3389/fmicb.2020.00858.
- Pertwee RG. “Emerging strategies for exploiting cannabinoid receptor agonists as medicines”. British Journal of Pharmacology 2009;156:397–411.
- Patricia H. Reggio “Endocannabinoid Binding to the Cannabinoid Receptors: What Is Known and What Remains Unknown”. Curr Med Chem. 2010; 17(14): 1468–1486.
- Katona L, Freund TF. “Endocannabinoid signaling as a synaptic circuit breaker in neurological disease”. Nat. Med. 2008;14:923–930.
- Adarsh Thomas Anthony, Shermeen Rahmat, Prerna Sangle, et.al. “Cannabinoid Receptors and Their Relationship With Chronic Pain: A Narrative Review”, Cureus. 2020 Sep; 12(9): e10436.
- Jason Socrates Bardi “Turning Off Pain’s Pathways”, The Scripps Research Institute Journal, Volume 1, Issue 22, August 13, 2001.
- David W. Self “Anandamide: a candidate neurotransmitter heads for the big leagues” Nature Neuroscience volume 2, pages303–304 (1999).
- Sabatino Maione, Fabiana Piscitelli, Luisa Gatta, Daniela Vita, et. al. “Non-psychoactive cannabinoids modulate the descending pathway of antinociception in anesthetized rats through several mechanisms of action”, British Journal of Pharmacology.
- Hill A. J., Jones N. A., Smith I., et al. “Voltage-gated sodium (NaV) channel blockade by plant cannabinoids does not confer anticonvulsant effects per se.” Neuroscience Letters . 2014;566:269–274.
- Ruhaak L. R., Felth J., Karlsson P. C., Rafter J. J., Verpoorte R., Bohlin L. “Evaluation of the cyclooxygenase inhibiting effects of six major cannabinoids isolated from Cannabis sativa.” Biological and Pharmaceutical Bulletin . 2011;34(5):774–778.
- Alhamoruni A., Wright K. L., Larvin M., O’Sullivan S. E. “Cannabinoids mediate opposing effects on inflammation-induced intestinal permeability.” British Journal of Pharmacology . 2012;165(8):2598–2610.
- Couch D. G., Maudslay H., Doleman B., Lund J. N., O’Sullivan S. E. “The use of cannabinoids in colitis: a systematic review and meta-analysis. Inflammatory Bowel Diseases”, 2018;24(4):680–697.
- Francesca Borrelli et al. “Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease”, Biochemical Pharmacology Volume 85, Issue 9, 1 May 2013, Pages 1306-1316.
- Giacoppo S., Gugliandolo A., Trubiani O., et al. “Cannabinoid CB2 receptors are involved in the protection of RAW264.7 macrophages against the oxidative stress: an in vitro study” European Journal of Histochemistry . 2017;61(1):p. 2749.
- Viviana di Giacomo et al. “Neuroprotective and Neuromodulatory Effects Induced by Cannabidiol and Cannabigerol in Rat Hypo-E22 cells and Isolated Hypothalamus” Antioxidants, 9(1), 71, 13 January 2020.
- K Wilson, M Auer, M Binnie, et al. “Overexpression of human kynurenine-3-monooxygenase protects against 3-hydroxykynurenine-mediated apoptosis through bidirectional nonlinear feedback” Cell Death Dis. 2016 Apr; 7(4): e2197.
- Delong Zhen, Junjun Liu, Xu Dong Zhang, and Zehua Song “Kynurenic Acid Acts as a Signaling Molecule Regulating Energy Expenditure and Is Closely Associated With Metabolic Diseases” Front Endocrinol (Lausanne). 2022; 13: 847611.
- Cathy Cassata, Allison Young “What is Norepinephrine?” American College of Lifestyle Medicine, February 13, 2023.
- Dolivo, D.M.; Larson, S.A.; Dominko, T. “Tryptophan metabolites kynurenine and serotonin regulate fibroblast activation and fibrosis.” Cell. Mol. Life Sci. 2018, 75, 3663–3681.
- Lin, H.M.; Barnett, M.P.; Roy, N.C.; Joyce, N.I.; Zhu, S.; Armstrong, K.; Helsby, N.A.; et al. “Metabolomic analysis identifies inflammatory and noninflammatory metabolic effects of genetic modification in a mouse model of Crohn’s disease. “ Journal of Proteome Res. 2010, 9, 1965–1975.
- Marciniak, S.; Wnorowski, et al., “Kynurenic Acid Protects against Thioacetamide-Induced Liver Injury in Rats.” Anal. Cell. Pathol. 2018, 2018, 1270483.
- Zheng, X.; Hu, M.; Zang, X.; Fan, et al. “Kynurenic acid/GPR35 axis restricts NLRP3 inflammasome activation and exacerbates colitis in mice with social stress.” Brain Behav. Immun. 2019, 79, 244–255.
- Parrott, J.M.; Redus, L.; O’Connor, J.C. “Kynurenine metabolic balance is disrupted in the hippocampus following peripheral lipopolysaccharide challenge.” Journal of Neuroinflammation 2016, 13, 124.
- Lutz B. “Endocannabinoid signals in the control of emotion.” Current Opinion in Pharmacology . 2009;9(1):46–52.
- Janero D. R., Makriyannis A. “Cannabinoid receptor antagonists: pharmacological opportunities, clinical experience, and translational prognosis.” Expert Opinion on Emerging Drugs . 2009;14(1):43–65.
- Cascio M. G., Gauson L. A., Stevenson L. A., Ross R. A., Pertwee R. G. “Evidence that the plant cannabinoid cannabigerol is a highly potent α2-adrenoceptor agonist and moderately potent 5HT1A receptor antagonist.” British Journal of Pharmacology . 2010;159(1):129–141.
- Rock E. M., Goodwin J. M., Limebeer C. L., et al. “Interaction between non-psychotropic cannabinoids in marihuana: effect of cannabigerol (CBG) on the anti-nausea or anti-emetic effects of cannabidiol (CBD) in rats and shrews.” Psychopharmacology (Berl) . 2011;215(3):505–512.
- De Vry J. “5-HT1A receptor agonists: recent developments and controversial issues.” Psychopharmacology (Berl) . 1995;121(1):1–26.
- Stahl SM, Gastpar M, Keppel Hesselink JM, Traber J (1992) “Serotonin~A receptors in depression mid anxiety.” Raven Press, New York.
- Zagzoog A., Mohamed K. A., Kim H. J. J., et al. “In vitro and in vivo pharmacological activity of minor cannabinoids isolated from Cannabis sativa.” Scientific Reports . 2020;10(1).
- Pucci M., Rapino C., Di Francesco A., Dainese E., D’Addario C., Maccarrone M. “Epigenetic control of skin differentiation genes by phytocannabinoids.” British Journal of Pharmacology . 2013;170(3):581–591.
- Dunn L. K., O’Neill J. L., Feldman S. R. “Acne in adolescents: quality of life, self-esteem, mood, and psychological disorders.” Dermatology Online Journal . 2011;17(1):p. 1.
- Oláh A., Markovics A., Szabó-Papp J., et al. “Differential effectiveness of selected non-psychotropic phytocannabinoids on human sebocyte functions implicates their introduction in dry/seborrhoeic skin and acne treatment.” Experimental Dermatology . 2016;25(9):701–707.
- Maccarrone M, Di Rienzo M, Battista N, Gasperi V et al. “The endocannabinoid system in human keratinocytes. Evidence that anandamide inhibits epidermal differentiation through CB1 receptor-dependent inhibition of protein kinase C, activation protein-1, and transglutaminase.” Journal of Biological Chemistry 278: 33896-903.
- Paradisi A, Pasquariello N, Barcaroli D, Maccarrone M (2008) “Anandamide regulates keratinocyte differentiation by inducing DNA methylation in a CB1 receptor-dependent manner.” Journal of Biological Chemistry 283: 6005-12.





